The following review of grass plants is intended to describe the functions of various plant parts that will be important in studying grass growth and regrowth.
Seeds
Grasses are herbaceous (nonwoody) plants with jointed stems, slender, sheathing leaves and flowers borne in spikelets. The grass family (Poaceae, formerly Gramineae) is divided into subfamilies. Most grasses in the United States are in the Pooideae, Panicoideae, and Chloridoideae subfamilies. Though grasses are herbaceous, with jointed stems, the categorization into subfamilies is made by more intricate plant anatomy.
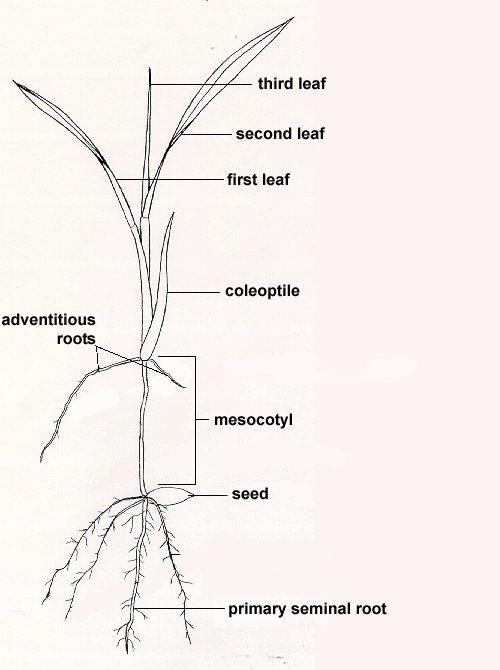
Grasses are monocotyledonous because the seeds contain only one cotyledon (seed leaf, also called the scutellum) (Fig. 1, corn kernel diagram). The coleoptile is enclosed in the cotyledon, a sheath which develops in the seed then breaks away and pushes upwards to the soil surface. This coleoptile is leaf-like in appearance and develops from the seed shortly after the radicle (root) appears (Fig. 2, coleoptile). In contrast, legumes, such as clovers, are dicotyledonous. Their seeds have two cotyledons. This difference means that grass seedlings emerge from the soil with only one leaf-like structure (Fig. 3, germination and emergence of monocots; Fig. 4, monocot). Legumes emerge from the soil with two cotyledons (Fig. 5, dicotyledons).
Roots
Initially, there are two root systems which support a grass seedling: 1) the seminal root which arises from the root itself and 2) the adventitious or secondary roots which arise from the crown node located at the base of the coleoptile. Seminal roots are called primary roots because they develop first. The seminal roots function until adventitious roots become established. There are no roots on the mesocotyl. The mesocotyl is an underground stem segment often called the subcrown internode or real stem. It elongates in a manner that helps push the coleoptile upward through the soil crust. This is important because the leaf enclosed within the coleoptile might unfurl beneath the soil surface and thus fail to emerge.
The first results of germination are the enlargement of the coleoptile and coleorhiza, a sheath that protects the primary root. This is followed by elongation of the primary root and the mesocotyl. Within days, additional mesocotyl roots develop. Roots developing in the crown area are called adventitious roots. Adventitious roots are the roots of the mature grass plant. The seminal roots will disappear. The roots anchor the plant to the ground, absorb nutrients and water from the soil, and function as carbohydrate storage tanks (Fig. 6, root diagram).
Crown
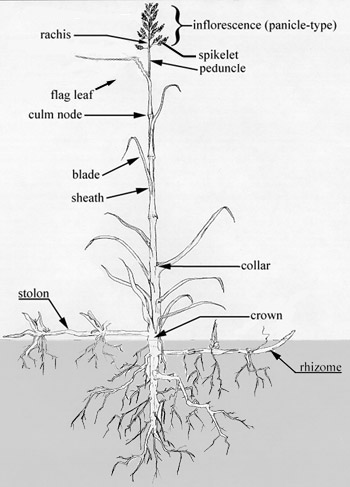
The crown is the base of the grass plant. It is the connecting tissue between the roots and the shoots. The crown produces buds that are the source of new tillers, adventitious roots, rhizomes, and stolons. This area is critical in understanding the regrowth of grass plants because it is the area to find buds which determine if regrowth will be successful (Fig. 7, crown). Some grass species have storage organs called corms that develop in the crown area (Fig. 8, corm; Fig. 9, corm at the base of the plant).
Stems
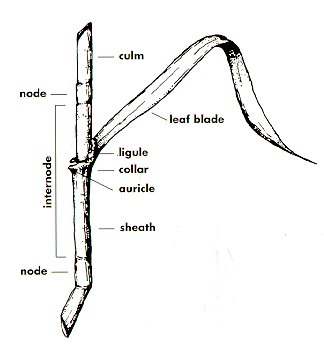
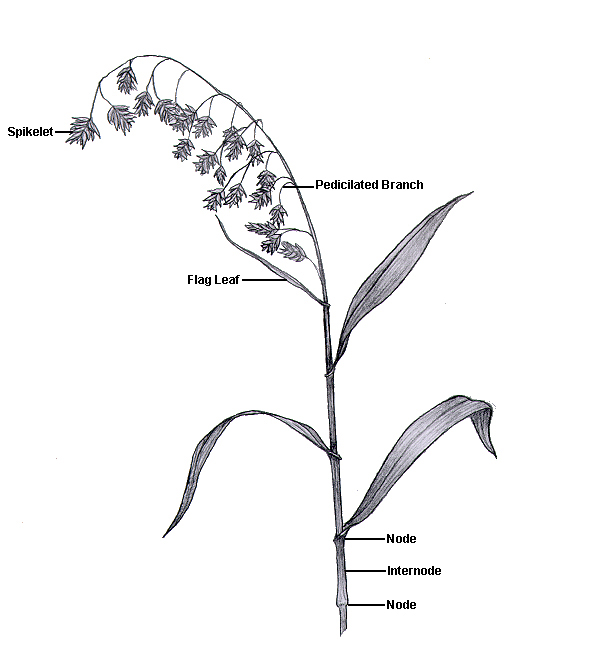
The flowering stem (culm) of grasses is comprised of nodes and internodes yielding a characteristic "jointed" stem (Fig. 10). Grass stems have solid joints at the nodes with hollow or pith-filled internodes. In contrast, rushes and sedges are without nodes and internodes and have a triangular stem shape (Fig. 11, common yellow sedge; Fig. 12, yellow nutsedge stem without nodes and internodes; Fig. 13, the tri-leaf formation of sedges).
Leaves
A grass leaf consists of a blade, collar, and supporting sheath (Fig. 14, Fig. 15).
Sheaths arise from each node. Blades are displayed alternately on opposite sides of the culm (Fig. 16).
Monocot leaf blades have parallel veination while dicots have netted veination (Fig. 17, grass veination; Fig. 18, dicot veination).
Species are often distinguished by variation in the size and shape of ligules and auricles, appendages located at the collar which joins the blade to the sheath (Fig. 19 and Fig. 20).
Inflorescences
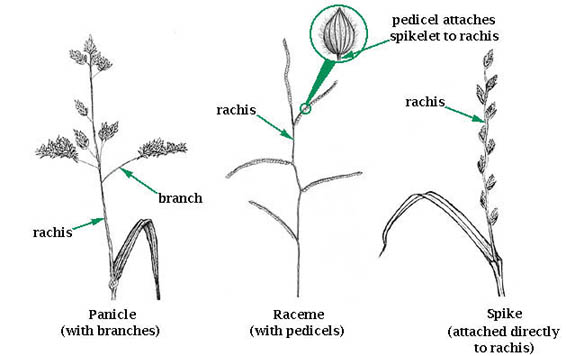
Grasses have three main inflorescence (seed head) types: panicle, spike, and raceme (Fig. 21). Each is unique as to how the individual flowering units, called spikelets, are attached to the central axis. The central axis of an inflorescence is called a rachis (Fig. 22).
Panicle inflorescences have spikelets individually supported by pedicels attached to panicle branches, not directly to the main axis (rachis) (Fig. 23). Panicles are the most common grass inflorescence but can have two forms: spreading and compact.
Spike inflorescences have spikelets that are sessile (attached directly to) the rachis without pedicels or branches. Spike inflorescences may exist in three forms:
For each of the above types, spikelets are attached directly to the rachis without pedicels or panicles branches.
Raceme inflorescences have spikelets born individually on short pedicels or stalks attached directly to the rachis. There are no branches. There are two forms.
Spikelet
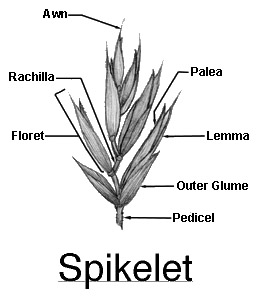
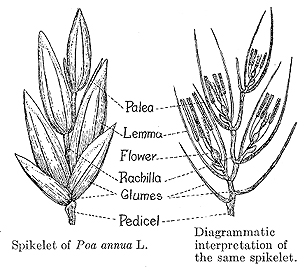
The most obvious unit of the grass inflorescence is the spikelet, comprised of a pair of glumes which enclose one or more florets. The number of florets per spikelet varies widely among the grass tribes (Fig. 32). This figure shows a pedicellate spikelet of the tall fescue panicle inflorescence. Notice that with multiple florets, each floret is born on a segmented central axis called the rachilla. In threshed form, each seed retains its rachilla segment (often called the rachilla joint). This segment is useful when identifying seeds (Fig. 33).
A floret is the reproductive unit of a spikelet.
In multifloreted spikelets, the florets are attached to a central axis called the rachilla. Figure 34 shows a floret (Fig. 34). Figure 35 shows a cross section of spikelets (Fig. 35).
At maturity, each floret (seed) of multi-floreted species retains its uniquely shaped rachilla segment. The unique shape and size of the rachilla segment is useful for identifying grass seeds (i.e. perennial vs. annual ryegrass; wedge-wise vs. cylindrical).
With single-floreted spikelets, like red top, reed canarygrass, meadow foxtail, and timothy, there is no rachilla and therefore, no rachilla segment on the threshed seeds as shown on figure 36, a meadow foxtail seed (Fig. 36).
Florets
The reproductive unit within a spikelet is called a floret. A floret is so named because it is a reduced (or modified) flower. It has no calyx or corolla as is found on most dicotyledonous flowers. The reproductive organs of the floret are enclosed by two bracts: the larger of the the two is called the lemma, the smaller is called the palea (Fig. 37). For cross pollination to occur, the lemma and palea must become separated. This separation occurs when spongy cells called lodicules, located in the base of the floret swell, due to the absorption of water spreading the floral bracts. Stamens within the floret extend, exposing the anthers to the wind and other spreading agents such as bees and insects (Fig. 38) .
This process of pollen shedding is called anthesis (Fig. 39).
Hannaway, D.B. 1998. Forage Identification CD-ROM. Oregon State University http://forages.orst.edu/resources/media/idcdrom/default.html
Chapman, C.P. 1998. The Biology of Grasses. CAB International, New York, NY.
Hitchcock, A.S.1950. Manual of Grasses of the United States. United States Government Printing Office. Washington, D.C.
Langer R.H.M. and G. D. Hill 1991. Agricultural Plants. Cambridge University Press, Cambridge, England.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}