Aphids feeding on phloem in the leaf blades and stems debilitate their host when populations are high. Tall fescue is a host to several species of aphids, including bird cherry-oat aphid [Rhopalosiphum padi (L.)] (Fig. 9-10), greenbug [Schizaphis graminum (Rondani)] (Fig. 9-11), Russian wheat aphid [Diuraphis noxia (Kurdjumov)], and yellow sugarcane aphid [Sipha flava (Forbes)]. Rhopalosiphum padi is present in Australia and New Zealand but is not considered to be a pest of tall fescue. In the United States aphids are minor pests of tall fescue but may cause severe damage in localized areas when environmental conditions are suitable. Certain biotypes of S. graminum may be more virulent on tall fescue than others (Kindler and Spomer, 1986). Festuca species, although less favorable hosts than cereals, were better hosts of D. noxia than other grasses, and infested plants exhibited stunted growth (Kindler and Springer, 1989; Kindler et al., 1991). Tall fescue harboring D. noxia in the field may act as a source for infestations on barley (Hordeum vulgare L.) and wheat (Triticum spp.), both of which can be damaged seriously by this pest. Schizaphis graminum can cause necrotic lesions when they inject salivary toxins into the leaf blade, whereas R. padi can transmit barley yellow dwarf virus to tall fescue. Almost 60% of tall fescue plants in a survey in Missouri were found to contain a virus transmitted by R. padi that produced symptoms in oat (Avena sativa L.) typical of barley yellow dwarf virus (Grafton et al., 1982). In their study of the aphid community on a variety of cool-season perennial grass accessions, Clement et al. (1990a) concluded that aphids had the potential to diminish plant health and seed production.
| Click to Expand | ||||
 |
 |
|
||
|
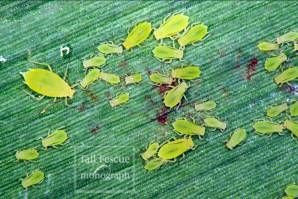
Fig. 9-10. Bird cherry-oat aphid, Rhopalosiphum padi. Left: adult (photo: Fred Baxendale); right: colonizing a leaf blade (photo: Steve Clement). |
Fig. 9-11. Greenbug, Schizaphis graminum (photo: Fred Baxendale).
|
Fig. 9-12. Root aphid, Aploneura lentisci. Left: root aphid; right: root aphid colonies (photos: AgResearch).
|
||
Aphids not only infest the foliage of tall fescue plants, but two species, Aploneura lentisci (Passerini) (Fig. 9-12) and Rhopalosiphum insertum (Walker), utilize roots of grasses as secondary hosts. Aploneura lentisci occurs in Europe and is widespread in New Zealand, where it inhabits roots of ryegrass and tall fescue all year. A very similar species has been observed on roots of tall fescue and ryegrass in Australia. Apple trees (Malus domestica Borkh.) are a primary host of R. insertum, which also is found throughout Europe, Australia, New Zealand, and the United States. Like A. lentisci, natural infestations of R. insertum have been found on tall fescue plants. Little is known about the economic importance of either species for their secondary grass hosts, but there is some evidence, in perennial ryegrass at least, that A. lentisci can reduce plant productivity (Popay and Gerard, 2007). Like its close relative R. padi, R. insertum also can transmit barley yellow dwarf virus (Halbert et al., 1992)
Numerous studies have been conducted on the sensitivity of aphids to endophyte in tall fescue and perennial ryegrass. Both R. padi and S. graminum prefer feeding on E- tillers over E+ tillers, and the survival and reproduction of both these species are reduced when they are confined to E+ plants (Johnson et al., 1985; Latch et al., 1985; Eichenseer et al., 1991; Eichenseer and Dahlman, 1992; Breen, 1993a). The novel nontoxic endophyte named MaxQ (United States; trademarked by Pennington Seed, Madison, GA) or MaxP (Australia and New Zealand; trademark of Grasslanz Technology Ltd., Palmerston North, New Zealand) also affects R. padi adversely (Bultman et al., 2006). In the field, E+ tall fescue is less likely to be infected by barley yellow dwarf virus, which is transmitted by R. padi (West et al., 1990; Mahmood et al., 1993). Diuraphis noxia shows a similar preference for E- tall fescue (Clement et al., 1990b; Springer and Kindler, 1990). Kindler et al. (1991) found that the survival of D. noxia nymphs confined to the base of E+ tall fescue turfgrass cultivars was much lower than on E- tillers, but survival of adult aphids was not affected. Feeding and survival of S. flava also were reduced by the presence of endophyte (Breen, 1993a). The corn leaf aphid [R. maidis (Fitch)], which can utilize tall fescue as a host but is not considered to be a pest of this plant, appears to be affected less by N. coenophialum than R. padi, while the English grain aphid [Sitobium avenae (F.)] showed no response to the presence of endophyte (Johnson et al., 1985). Latch et al. (1985) observed that endophyte infection in tall fescue did not alter the behavior of the rose-grain aphid [Metalophium dirhodum (Walker)] or the grain aphid [Sitobium fragariae (Walker)]. They also showed that infection of tall fescue by a Phialophora-like endophyte did not inhibit feeding of R. padi and that these aphids, in a choice test, actually preferred plants infected with this endophyte.
Nontoxic endophytes, such as MaxQ, reduced populations of A. lentisci on tall fescue (Jensen and Popay 2007). A similar effect has been observed recently with R. insertum on roots of tall fescue, where numbers on E- plants averaged 86/plant, while those on plants infected with MaxQ averaged 0.3/plant (P < 0.001) (A.J. Popay, unpublished data, 2008).
The effect of tall fescue endophyte on aphids has been linked with the production of loline alkaloids. Johnson et al. (1985) demonstrated that the deterrent and toxic factors affecting R. padi were associated primarily with the methanol extract, which contained high concentrations of these pyrrolizidine alkaloids. Trials using a number of different plant-endophyte combinations with a range of alkaloid profiles confirmed the sensitivity of R. padi only to those plants infected with endophytes where lolines were produced (Siegel et al., 1990). Survival of S. graminum, however, was reduced on plant material containing peramine only, or on plants containing both peramine and lolines (Siegel et al., 1990). Using full-sibling fungal progeny of Epichloë festucae Leuchtm., Schardl, and Siegel, loline alkaloid expression cosegregated with activity against both R. padi and S. graminum,, and in planta levels of loline alkaloids correlated with anti-aphid activity (Wilkinson et al., 2000). Rhopalosiphum padi may be able to survive on E+ tall fescue where there is senescing leaf material because this tissue contains lower levels of loline alkaloids than younger leaves (Eichenseer et al., 1991).
| <--Previous | Back to Top | Next--> |
|
|
|
Oregon State University
Corvallis, OR 97331 Send Email |
