The endophytic Neotyphodium species, which include N. coenophialum and N. lolii and their close relatives the Epichloë species (Ascomycotina, Clavicipitaceae, Balansieae) (White, 1997), are biotrophs that form stable symbioses with grasses in the subfamily Pooideae (Schardl and Phillips, 1997; Schardl et al., 2004).
Neotyphodium endophytes (like most fungi) consist of elongate hyphal filaments (Fig. 14-1; see the enclosed CD for color versions of many of the figures in this chapter, which have better resolution than the black and white figures in the book). Nutrients from the host plant are absorbed directly through the hyphal wall. The hyphae are divided into compartments by septa, and compartments are interconnected by pores (Fig. 14-2) in the septa (Fig. 14-3), through which cytoplasm, including organelles, may pass.
| Click to Expand | ||||||||
|
|
 |
 |
||||||
|
Fig. 14-1. Hypha of Neotyphodium coenophialum (-->) in a tall fescue leaf sheath. |
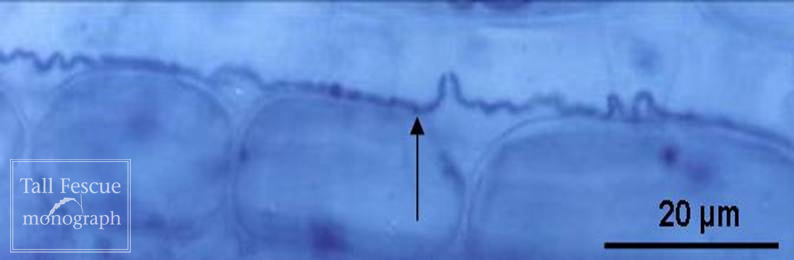
Fig. 14-2. Hypha of Neotyphodium coenophialum with septum (-->) in young leaf tissue. |
Fig. 14-3. Hypha (Epichloë typhina) with developing septum (-->) in perennial ryegrass stroma. |
||||||
Septal pores can be plugged by spherical Woronin bodies (Fig. 14-4), specialized peroxisomes that prevent cytoplasmic loss when hyphae are damaged (Tenney et al., 2000). Endophytes of tall fescue and other cool-season grasses are potent producers of alkaloids, such as peramine (Tanaka et al., 2005), ergovaline (Fleetwood et al., 2007; Schardl et al., 2006), lolitrem B (Young et al., 2006), and lolines (Schardl et al., 2007), and are predicted to produce more secondary metabolites than those currently characterized since they contain numerous genes implicated in the production of bioactives (Johnson et al., 2007).
For further reading on fungal morphology and morphogenesis see Moore (1998). For further reading on Neotyphodium and Epichloë refer to Christensen et al. (2002), Clay and Schardl (2002), Hinton and Bacon (1985), Koga et al. (1993), Philipson and Christey (1986), Schardl and Phillips (1997), and Siegel et al. (1987).
| Click to Expand |
 |
|
Fig. 14-4. Hyphal septum of Neotyphodium lolii in ryegrass. The septal pore in each compartment is bordered by a Woronin body (w). |
| <--Previous | Back to Top | Next--> |
|
|
|
Oregon State University
Corvallis, OR 97331 Send Email |